信号通路|荟萃9条最常见的信号通路( 三 )
所以这个时候glucose更多的是用来转化成氨基酸而非能量储存,所以mTOR未被激活 。相反在氨基酸富余而glucose不足的情况下,合成所需的能量补给不足,氨基酸这个时候更多的可能会被用来分解而提供细胞活动所需能量,所以mTOR也必然不能被激活 。
这正看出mTOR的调节模式也反映了细胞代谢平衡机智的策略 。比较有意思的是,两条线路都是经由小G蛋白的信号,那么细胞的代谢稳态比如氨基酸和糖类之间的生化转化的信号如何感知呢?是否小G蛋白参与其中起到重要的作用呢?引人遐想 。
2.3、对自噬的影响
mTOR对自噬的调节本质上也是生长和代谢之间的调节 。这其中有营养因素,也有能量因素 。自噬是当细胞的营养物质或能量不足时,细胞为了维持自己的基本生存需要,就会通过溶酶体降解一些相对次要百思特网的蛋白,以及一些相对多余的细胞器,来供给物质和能量 。
除了能量的主要载体ATP相对不足,即AMP/ATP上升,营养供应不足,mTORC1的活性也会不足,于是mTORC1释放了对自噬起始调节的抑制 。另一方面来自PI3K-AKT,MAPK cascades信号对mTORC1的调控也体现了生长调节与代谢之间的联系 。此外,P53/基因组的稳定性也可直接调节mTORC1的活性,也增加了这方面的证据 。
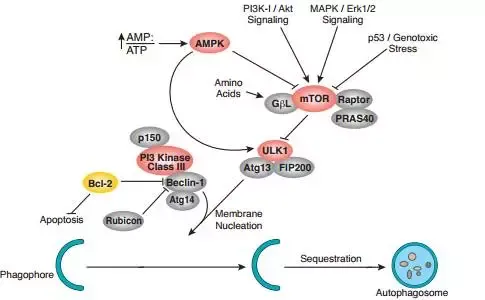
2.4、对肿瘤治疗的影响
既然mTOR对代谢有这么大的影响,众所周知,肿瘤就是个能耗大户,那抑制mTOR通路是不是就能影响肿瘤的生长呢?
还真有这种研究,雷帕霉素就是经典的mTOR的抑制剂,能够同时抑制住通路中两个大块头mTORC1和mTORC2的活性 。那是不是只要用它就能很好的限制住肿瘤呢?
没那么简单,下面的一百思特网个负反馈的图就很好的说明了这个原因 。
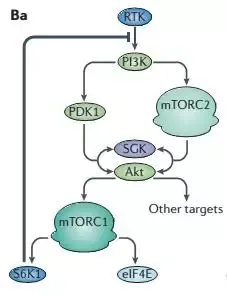
当mTOR被长期抑制后,S6K1的活性也会降低,S6K1对RTK的反馈抑制回路就会被解除,那么,将会有更多的补偿回路来补偿AKT和AGKs这些促进生存的因子,此外S6K1还可以抑制MEK-ERK信号通路,同样雷帕霉素可以解除对MEK-ERK的抑制,抑制临床上表现为雷帕霉素抵抗性 。
有研究表明,雷帕霉素并非完全抑制了mTORC1的全部活性,在不同的底物可能各有差别 。对于4E-BP1只是短暂或部分抑制其磷酸化,当S6K1-IRS1的负反馈回路使得PI3K-AKT的激活,最终可能还会导致4E-BP1的过磷酸化以及mTORC1其他促进肿瘤的功能 。
科学家试图设计mTOR的抑制剂以期对抗肿瘤 。竞争ATP结合的抑制剂Torin 1和PP242都有很好的临床疗效,但仍然无法回避S6K1-IRS1的反馈回路 。再想下去,如果保持mTORC1的活性保持对AKT活性负反馈的完整性,而只抑制mTORC2的活性 。
这样就可以抑制AKT的Ser473的磷酸化抑制其活性,然而后续的研究却发现,虽然AKT的Ser473的磷酸化被抑制,理论上Thr308的磷酸化不会被引发,实际的情况却是在此背景下Thr308位可能通过其他的路径得以补偿,mTORC2的抑制对AKT活性的影响恐怕只会影响到部分底物如FOXO1和FOXO3 。
而最近几年的策略集中在对mTOR和PI3K双重抑制剂,这两种蛋白激酶都属于PI3K蛋白激酶家族 。从原理上说,它不仅抑制了mTOR的活性,也直接抑制了AKT的活性,所以很好的规避了S6K1-IRS1的反馈回路 。双重抑制剂PI-103也展示了良好的杀死淋巴癌的效果,但是这样的双重抑制剂对正常的淋巴组织却一样有毒副作用 。针对mTOR的肿瘤治疗方案依然任重道远 。
对mTOR靶向治疗肿瘤的尝试也很好的证实肿瘤是一个种调动了整个系统的系统性疾病,这大概是肿瘤难以对付的最重要的原因 。对于简单的病因的肿瘤,治疗则相对较为容易 。
对于复杂的病因引起的肿瘤,可能最初也是由单因素引起的细胞内部对生长增殖自我约束机制的破坏,而简单点的破坏可能会被自我修复,也可能引起连锁的正反馈的约束的逃逸,引发肿瘤 。这好比细胞社会中,如果机体受到的外界因素的伤害不能抵御时,那么细胞个体便会不依赖整体协作而作为个体而努力存活下去 。
3. MAPK信号通路
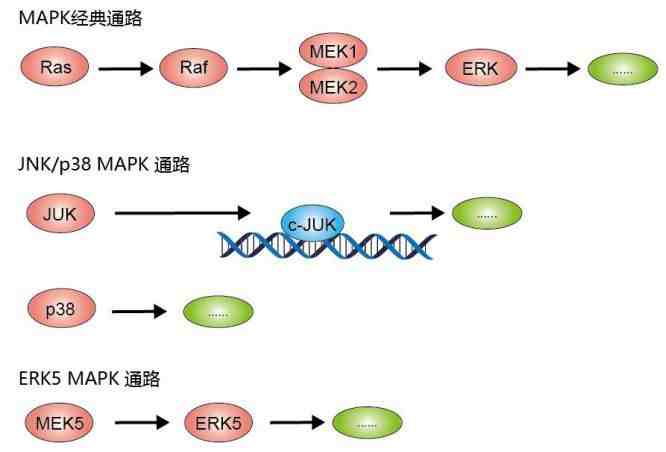
(图片截取于优秀学员Ewen的整理笔记)
MAPK(丝裂原活化蛋白激酶)通路有三级的信号传递过程:MAPK,MAPK激酶(MEK或MKK)以及MAPK激酶的激酶(MEKK或MKKK) 。这三种激酶能依次激活,共同调节着细胞的生长、分化、应激、炎症反应等多种重要的生理/病理效应 。
- 男人只是在耽误你的信号,趁早远离
- 教师节活动的新闻稿如何写?
- 教师节活动的新闻稿怎么写?
- 路由器价格表_无线路由器多少钱一个
- 输入微信号查询聊天记录软件「远程查看微信聊天记录软件」
- 古今诗词名句
- 胸部b超多少钱做一次_乳房癌症的早期信号
- 股票买入口诀,股票买入十大信号图解
- 齐齐哈尔贷款可以通过微信吗 「微信号」
- 苹果11怎么样,目前哪款苹果最值得买